



Hay tres aspectos de la planta que afectan la transpiracion:
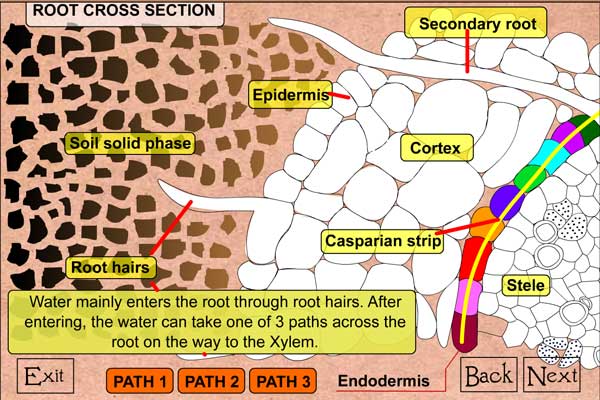
Detalles de la Raíz- La principal ruta de entrada de agua a las plantas es del suelo a las raíces. El agua entra por la región cercana a la punta de la raíz en crecimiento; la misma región donde crecen los pelos radicales. La superficie de los pelos radicales necesita estar en contacto con el suelo para acceder al agua ahí contenida. El agua se difunde al interior de la raíz, donde puede tomar al menos tres diferentes rutas para eventualmente llegar al xilema, que es el conducto que lleva el agua hasta las hojas. Pase al otro nivel de la animación para ver las rutas que el agua puede tomar en la raíz.
 |
| Corte transversal de la raiz de una planta. |
¿Cuál ruta toma el agua para ir de los pelos radicales hacia las hojas?
Una vez que el agua a entrado a un pelo radical, ésta debe moverse a través del cortex y la endodermis, antes de llegar al xilema. Corte transversal de la raíz de una planta El agua tomará la ruta de menor resistencia hacia el xilema, pudiendo moverse por tres diferentes vías. Una vía es la ruta apoplástica, donde el agua se mueve a través de las paredes celulares sin cruzar membrana alguna; es decir, el agua no entra a las células. Las otras dos vías, llamadas rutas celulares, requieren que las moléculas de agua crucen las membranas celulares.
La primera vía celular es la ruta transmembranal, por la cual el agua se mueve atravesando membranas para dejar una célula y entrar a otra. La segunda vía celular es la ruta simplástica en las que las moléculas de agua pasan de una célula a otra a través de las interconexiones celulares conocidas como plasmodesmos. Independientemente de la ruta que las moléculas de agua tomen, una vez que atraviesan el córtex deben ahora cruzar la endodermis. La endodermis es un anillo de células con una cubierta endurecida o cerosa conocida como banda de Caspari, la cual detiene el movimiento del agua entre células.
En este punto, el agua es forzada a través de las membranas de las células endodérmicas, creándose un efecto de coladera. Una vez que han atravesado la endodermis, las moléculas de agua entran libremente al xilema, donde se unen a la columna de agua, también conocida como torrente transpiratorio, que se dirige hacia las hojas.
Detalles del Xilema– El xilema es probablemente la parte más larga de la ruta del agua hacia las hojas. Es también la parte de la ruta que ofrece la menor resistencia al paso del agua, siendo alrededor de 1000 millones de veces menor que la resistencia al transporte del agua de célula a célula que involucra membranas. El xilema está compuesto por traqueidas y elementos de los vasos; los diámetros son menores en las primeras.
En ambos casos, las paredes celulares contienen celulosa y lignina; lo que las hace extremadamente rígidas. Las células del xilema no contienen membranas y son consideradas células muertas. En este tejido conductor no hay columnas simples de acarreo de agua; las células se encuentran sobrepuestas, creando una serie de vías alternas que puede tomar el agua conforme sube hacia las hojas.
|
Las plantas son más susceptibles a la cavitación cuando las tasas de transpiración son extremadamente altas. |
¿Cómo se abren los estomas? Los estomas detectan las condiciones ambientales, como la luz, para abrirse. Esos cambios desencadenan una serie de reacciones que ocasionan que las células oclusivas se llenen de agua. Para ilustrar lo anterior, sigamos los eventos que sucederían cuando amanece y el sol empieza a ascender, de forma que una planta de algodón recibe la señal para abrir los estomas.
|
Cavitación– La cavitación es el llenado de una traqueida o elemento del vaso con aire. También se le conoce como “embolismo”. Recordemos que durante la transpiración la columna de agua es jalada fuera de la planta por efecto de la evaporación en la superficie de las hojas; cuando la fuerza de tensión que jala al agua es mayor que la fuerza que mantiene a las moléculas de agua unidas, la columna de agua se rompe.
Usando sensores acústicos es posible oír el ruido que se produce cuando las moléculas de agua se separan. Las características estructurales únicas del xilema ayudan a la planta a confinar las burbujas de aire de modo que no interrumpan completamente el movimiento ascendente del agua.
Las plantas son particularmente sensibles a la cavitación durante las horas más calurosas del día, cuando no hay suficiente agua disponible en el suelo para mantener la alta demanda creada al aumentar la evaporación en la superficie de las hojas. Por otra parte, la cavitación también puede ocurrir a temperaturas congelantes. Debido a que la solubilidad del gas en el hielo es muy baja, el gas es liberado de la solución cuando ésta se congela en el xilema. El congelamiento de la solución del xilema es un problema sobretodo durante la primavera, cuando el hielo se derrite, dejando burbujas en los elementos del vaso. Estas burbujas pueden bloquear el transporte del agua y causar estrés hídrico en las hojas.
Las plantas evitan la cavitación o minimizan los daños mediante diferentes mecanismos:
- Las células del xilema poseen punteaduras o pequeños orificios que permiten el flujo del agua líquida, pero no permiten el escape de las burbujas de gas; esta característica estructural ayuda a mantener las burbujas de gas en una sola célula, de forma que otras células del tejido mantienen el transporte ascendente del agua.
- Las punteaduras facilitan el desvío del agua hacia otras células que no contienen burbujas de gas.
- Las burbujas de gas se disuelven nuevamente en el agua al disminuir la tensión que jala el agua en le xilema; por ejemplo, durante la noche, cuando el agua no está en ascenso por el cierre de los estomas.
- Las células del xilema con diámetros menores (traqueidas), comparadas con aquellas de diámetros más amplios (elementos del vaso), evitan la cavitación ya que la columna de agua es más resistente a la ruptura y la formación de burbujas.
Detalles de los Estomas– Los estomas son el principal mecanismo de control de las plantas para reducir la pérdida de agua y su capacidad de respuesta es muy rápida. Los estomas son sensibles a los cambios ambientales, lo que estimula la apertura o cierre estomático.
La principal función de los estomas es permitir la entrada de bióxido de carbono para la fotosíntesis, al tiempo que se permite la salida del agua conforme ésta se evapora de las hojas para enfriarlas. Dos células especializadas llamadas “células guarda” u oclusivas conforman cada estoma. Las plantas tienen una gran cantidad de estomas en las hojas, llegando a contarse hasta 400 estomas/mm2 en algunas especies. Algunas plantas presentan estomas en ambos lados de las hojas y otras solo en la parte inferior para minimizar la pérdida de agua.
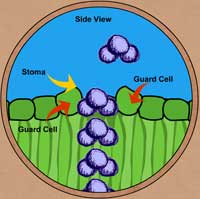 |
| Diagrama de moléculas de agua saliendo del estoma – vista lateral. |
VISTA LATERAL DE UN ESTOMA– Las condiciones ambientales que afectan la apertura y cierre estomáticos son la luz, agua, temperatura y la concentración de CO2 dentro de la hoja. Los estomas se abrirán en la luz y se cerrarán en la obscuridad; sin embargo, los estomas pueden cerrarse a medio día si el agua es limitante, si el, CO2 se acumula en las hojas o si la temperatura es muy alta. Si le falta agua a la planta, los estomas se cerrarán porque no habrá agua suficiente para crear la presión necesaria en las células oclusivas para que ocurra la apertura estomática; esta respuesta ayuda a la planta a conservar agua.
Si la concentración interna de CO2 se incrementa, los estomas reciben la señal para cerrarse ya que la respiración está liberando más CO2 del que está siendo usado en la fotosíntesis. Por lo tanto, no hay necesidad de mantener los estomas abiertos y perder agua si la fotosíntesis no está funcionando. Por el contrario, si el CO2 de las hojas se encuentra bajo, los estomas permanecerán abiertos para mantener su abastecimiento para la fotosíntesis.
Las altas temperaturas también sirven de señal para cerrar los estomas. Las altas temperaturas incrementan la pérdida de agua; con menos agua disponible, las células oclusivas se vuelven flácidas y el estoma se cierra. Otro efecto de las altas temperaturas es que las tasas respiratorias rebasan a las tasas fotosintéticas, causando un incremento en la concentración de CO2 en las hojas; esto causará también el cierre de los estomas. Recordemos que algunas plantas abren sus estomas bajo altas temperaturas para que la transpiración enfríe las hojas.
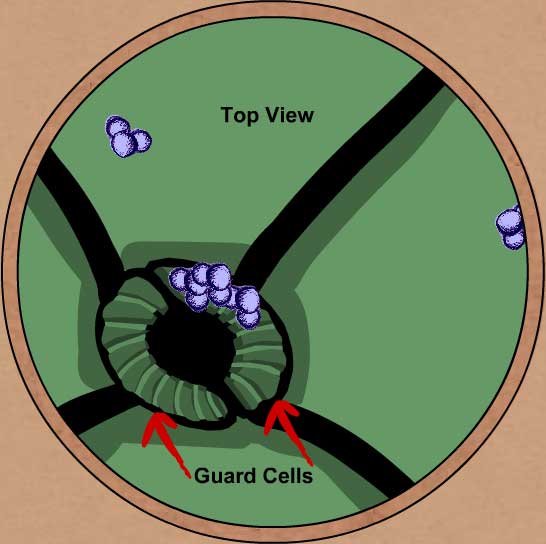
VISTA SUPERIOR DE LOS ESTOMAS
|
Las microfibrillas de celulosa evitan que las células oclusivas se expandan de manera uniforme y adquieran una forma esférica; la gruesa pared celular alrededor del poro y las microfibrillas dispuestas en forma radial al mismo restringen la expansión solo a la parte externa de las células. |
|
Muchas plantas presentan estomas únicamente en la parte interior de las hojas para ayudar a disminuir la pérdida de agua. |

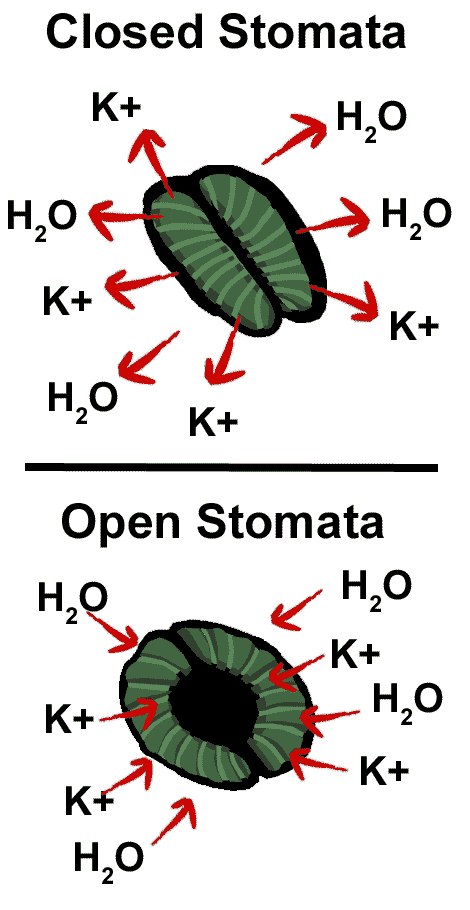
ESTOMAS ABIERTOS– Cuando los estomas reciben una señal para abrirse, iones potasio (K+) entran a la células oclusivas. Esto induce la entrada de agua siguiendo el diferencial de potencial hídrico, creando una presión hidrostática en estas células que cambia su forma. Cada célula oclusiva del par se expande en el lado exterior del estoma, pero no en el lado interno, adquiriendo una forma de riñón y formándose entonces la apertura o poro entre ambas células para que ocurra el intercambio gaseoso. Las células en forma de riñón son características de las dicotiledóneas; en otras plantas (por ejemplo, los pastos y otras monocotiledóneas) estas células presentan la forma de una pesa para ejercicios. La forma que adquieren las células oclusivas se debe a la presencia de microfibrillas de celulosa que rodean a la célula en forma radial al poro; algo parecido a las cuerdas de las llantas radiales. Estas microfibrillas son rígidas y no se extienden con la entrada de agua a la célula; la pared celular en la parte que rodea al poro estomático es más gruesa que en el resto de la célula, evitando que ese lado se expanda. Por lo tanto, cuando la presión interna de la célula aumenta por la entrada de agua, las células oclusivas no se hacen más amplias sino que los lados externos se expanden en forma desproporcionada a los lados internos; esta expansión desigual permite la formación del poro entre ambas células oclusivas.
ESTOMAS CERRADOS– Los estomas deben abrirse para que la fotosíntesis ocurra. Sin embargo, con los estomas abiertos existe el riesgo de perder mucha agua por transpiración. Los estomas se cierran cuando las células oclusivas se vuelven flácidas por la pérdida de agua y presión interna. Esto ocurre debido a que los iones potasio (K+) salen de las células oclusivas y son seguidos por el agua, disminuyendo entonces la presión en estas células.