



Introducción
La variación genética originada en cualquier especie por procesos como mutaciones, recombinación, hibridación interespecífica, introgresión, transformación genética, selección o deriva genética, puede ser asociada con zonas geográficas bien definidas en lo que se denomina centros de origen o centros de diversidad. La dinámica de esta variación en las distintas especies ha sido modulada por la forma en que se da su reproducción, es decir, por los mecanismos que permiten, en cada una de las especies la transmisión de los caracteres que conforman su identidad. Los distintos sistemas de reproducción van a originar distintos niveles de homocigosis o heterocigosis en los genes que conforman al individuo, y por supuesto que conforman la población. Esto es lo que se denomina estructura genética de una población, concepto de gran utilidad dentro de un programa de mejoramiento genético, ya que de él va a depender las estrategias a utilizar para este fin.
 1) Tipos de reproducción
1) Tipos de reproducción
Existen dos formas de reproducción en las plantas: la sexual y la asexual. En la primera debe existir la formación de gametos (masculinos y femeninos) los cuales al unirse formarán el nuevo individuo. Esos gametos provienen del proceso de división celular reduccional llamado meiosis, en el cual, a partir de una célula somática se obtienen células sexuales con la mitad de la carga cromosómica de la célula madre. En la reproducción asexual, tan solo se da el proceso de división celular denominado mitosis, en el cual, a partir de una célula somática se obtienen dos células con la misma carga cromosómica de la célula madre. Esta situación permite explicar la diferencia en variabilidad genética entre especies de reproducción sexual y especies de reproducción asexual, en las primeras hay una gran variabilidad mientras que las segundas son prácticamente invariables.
 Dentro de las especies de reproducción sexual, existen dos formas en las cual este se puede dar, dependiendo de la procedencia del gameto masculino que fecunda al gameto femenino, es decir, dependiendo de la forma como se da la polinización. Cuando el gameto masculino proviene de la misma flor que porta el gameto femenino, se hace referencia a la autogamia o autopolinización. Por supuesto, esto sólo puede ocurrir en especies de flores perfectas. Por el contrario, cuando el gameto masculino proviene de una planta distinta a la que porta el gameto femenino, se hace referencia a la alogamia o polinización cruzada, este proceso puede darse en especies con flores perfectas o flores unisexuales. La autogamia y la alogamia van a dar origen a poblaciones con una estructura genética distinta.
Dentro de las especies de reproducción sexual, existen dos formas en las cual este se puede dar, dependiendo de la procedencia del gameto masculino que fecunda al gameto femenino, es decir, dependiendo de la forma como se da la polinización. Cuando el gameto masculino proviene de la misma flor que porta el gameto femenino, se hace referencia a la autogamia o autopolinización. Por supuesto, esto sólo puede ocurrir en especies de flores perfectas. Por el contrario, cuando el gameto masculino proviene de una planta distinta a la que porta el gameto femenino, se hace referencia a la alogamia o polinización cruzada, este proceso puede darse en especies con flores perfectas o flores unisexuales. La autogamia y la alogamia van a dar origen a poblaciones con una estructura genética distinta.
Dentro de las especies de reproducción asexual, se pueden distinguir la apomixis vegetativa y la agamospermia como formas de reproducción, sin embargo, por no tener consecuencias distintas sobre la estructura genética de la población estas no serán discutidas.
2) Mecanismos de control de la polinización
La autogamia y la alogamia son extremos de una escala de variación continua, es decir, normalmente no se encuentran especies totalmente autógamas o totalmente alógamas, sino que hay cierto nivel de ambos. Por esta razón, según el predominio de alguna de estas situaciones, se hace referencia a plantas predominantemente autógamas y plantas predominantmente alógamas.
2.1. Mecanismos que favorecen la autogamia
Cleistogamia es la situación en la cual las flores nunca dejan descubiertos sus órganos sexuales, es decir, los pétalos siempre mantienen cubiertos al androceo y al gineceo, esto permite que la fecundación se realice necesariamente entre gametos de una misma flor. La cleistogamia es muy común en varias especies de leguminosas. En otras especies como el ajonjolí, la apertura de la flor es posterior a la fecundación, por tal razón esto se ha llamado cleistogamia efectiva. Otro caso de cleistogamia efectiva se presenta en tomate, donde aún después de la apertura de la flor, los estambres se disponen de tal manera que cubren completamente el gineceo, por lo cual en esta especie también se favorece la autogamia. Entre algunas especies autógamas de importancia económica se pueden mencionar la caraota, el frijol, quinchoncho, ajonjolí, tomate, pimentón, arroz, trigo, papa, soya y tabaco.
2.2. Mecanismos que favorecen la alogamia
 2.2.1. Separación de gametos en el espacio
2.2.1. Separación de gametos en el espacio
a) Monoecia: existen algunas especies vegetales que poseen flores unisexuales las cuales, tanto masculinas como femeninas, se hallan sobre una misma planta como es el caso del maíz. En estas especies es más probable que el polen emitido por una planta fecunde óvulos de otra planta, es por esto que se considera la monoecia como un mecanismo que favorece la alogamia.
b) Dioecia: cuando una especie posee flores unisexuales, pero cada uno de los sexos se halla sobre distintas plantas, se hace referencia a la dioecia, Indudablemente, la única forma de descendencia en este tipo de especies es mediante la alogamia. Esto ocurre en el espárrago.
2.2.2. Separación de gametos en el tiempo
Una forma efectiva que tienen algunas especies de favorecer la alogamia es la dicogamia, que nos es mas que la madurez diferencial de los órganos sexuales masculinos y femeninos, es decir, cuando uno de ellos se encuentra sexualmente maduro, produciendo gametos, el otro aún está inmaduro, y cuando empieza a formar gametos ya el otro ha perdido su funcionalidad. Esto ocurre en aguacate y en el maíz.
2.2.3. Incompatibilidad
En algunas especies vegetales tales como el tabaco, al momento del contacto de un grano de polen con el estigma de la misma flor se da una reacción bioquímica que conduce al necrosamiento del estigma y por tanto a una falla en la polinización, por esta razón la incompatibilidad induce a que la descendencia provenga de polinización cruzada. Existen dos mecanismos que permiten la incompatibilidad, el sistema gametofítico y el esporofítico, esto dependiendo del genotipo responsable de la reacción de incompatibilidad, si el genotipo del grano de polen o el genotipo de la planta que originó el grano de polen.
a) Sistema gametofítico: el genotipo haploide del grano de polen será el responsable de la reacción de incompatibilidad. Esta se da cuando gen que determina el genotipo del grano de polen es igual a uno de los dos alelos presentes en las células del estigma (figura 1).
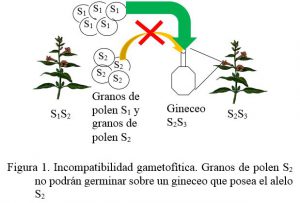
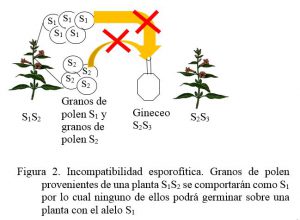
b) Sistema esporofítico: el genotipo del esporofito, es decir de la planta madre que da origen a los granos de polen, determina la reacción de incompatibilidad. Esta racción la provoca un genotipo diploide, se establece la interacción intraalélica de dominancia. Generalmente en estos genes se presentan series de alelos múltiples. Esto determina un efecto materno en el cual los granos de polen no se comportarán como lo dicta su propio genotipo, sino como el alelo dominante presente en su planta madre. Cuando este coincide con alguno de los alelos presentes en las células del estigma, se da la incompatibilidad (Figura 2).
 Los mecanismos que favorecen la alogamia, sea cual sea, lograrán que los granos de polen transportados por el viento o por insectos puedan germinar en gineceos de otras plantas, no de la planta en donde se dio su producción. La polinización que se da por insectos es denominada polinización entomófila, mientras que la que se da por el viento se denomina polinización anemófila.
Los mecanismos que favorecen la alogamia, sea cual sea, lograrán que los granos de polen transportados por el viento o por insectos puedan germinar en gineceos de otras plantas, no de la planta en donde se dio su producción. La polinización que se da por insectos es denominada polinización entomófila, mientras que la que se da por el viento se denomina polinización anemófila.
2.2.4. Androesterilidad
En forma espontánea se presentan en la naturaleza individuos incapaces de producir polen, o si lo producen este es inviable. Esta situación ha sido aprovechada agronómicamente en la producción de híbridos. Dependiendo del ADN que determine esta característica, la androesterilidad es clasificada como génica, citoplasmática y génica – citoplasmática.
a) Androesterilidad génica: este tipo de androesterilidad está condicionado por el alelo nuclear recesivo ms , que en condición homocigota impide la producción de polen. Debido a que el genotipo msms no puede producir polen, pero es valioso agronómicamente para facilitar la producción de híbridos, hay la necesidad de multiplicar este material con individuos fértiles heterocigóticos Msms. Esto trae como consecuencia que descendencia sea 50 % fértiles y 50 % estériles, lo cual conlleva a un trabajo adicional en lo que se refiere a la eliminación manual de las plantas fértiles. Esto representa una seria desventaja desde el punto de vista económico, sin embargo es ventajoso en el sentido que los individuos híbridos obtenidos después del esntresaque manual sean 100 % fértiles.
b) Androesterilidad citoplasmática: en las mitocondrias y en los cloroplastos existe ADN que puede codificar alguna proteína que impida la formación del polen. Estando ambos organelos en el citoplasma, y conociendo que la herencia de ellos se da por la vía materna, es de esperar que cualquier cruzamiento con individuos con androesterilidad citoplasmática va a resultar en una descendencia totalmente androesteril. Esta situación es favorable para el mantenimiento de la semilla estéril, pero desfavorable para la producción de híbridos en especies cuya producción comercial es su semilla, ya que por ser estéril restringe su uso a especies cuyo producto comercial sea vegetativo, como ocurre en cebolla, lechuga, repollo, papa, etc.
c) Androesterilidad génica – citoplasmática: este tipo de androesterilidad se da cuando genes nucleares y citoplasmáticos interactúan para originarla. La única combinación
posible entre esos genes para propiciar la androesterilidad es que en ambos compartimientos celulares exista la información para androesterilidad. Basado en esto se presentan tres tipos de líneas en la explotación de este sistema:
Línea A o androesteril, denotada como [ E ] msms
Línea B o mantenedora denotada como [ F ] msms
Línea R o restauradora denotada como [ F ] MsMs
La líneas A es multiplicada al cruzarla con la línea B, dando una descendencia 100 % estéril. Esta condición es ideal para la producción de híbridos, superando la desventaja que en este sentido tiene la androesterilidad génica. El cruzamiento de la línea A con la línea R resultará en el híbrido deseado, el cual tendrá el 100 % de sus individuos fértiles, superando la desventaja de la androesterilidad citoplasmática.
3) Cuantificación del nivel de polinización cruzada
Como se mencionó anteriormente, las especies vegetales poseen cierto nivel de autogamia y cierto nivel de alogamia. Para determinar el porcentaje de polinización cruzada que se presenta en una especie, se pueden sembrar dos hileras de plantas que difieran en una característica fácilmente observable y de herencia sencilla y conocida. Si se cosecha un número determinado de plantas con el carácter recesivo, y se siembra su semilla, puede fácilmente determinarse que semillas provienen de autopolinizaciones (las que presenten el carácter recesivo) y que semillas provienen de polinización cruzada (las que presenten el carácter dominante). Con esta proporción se habrá determinado el nivel de polinización cruzada.
4) Estructura genética de las poblaciones
La estructura genética de las poblaciones de plantas de reproducción asexual dependerá exclusivamente del genotipo del material que le dio origen. Si se considera sólo un gen, al ser el material original homocigoto para ese gen, la población que de allí se origine será igualmente homocigota, mientras que si es heterocigoto así será la población.
En plantas autógamas, al considerar solo un gen, se puede observar (cuadro 1) como cada generación de autogamia disminuye en un 50 % la heterocigosis, resultando después de n generaciones de autofecundación, poblaciones virtualmente homocigotas.
En plantas alógamas, se cumple el equilibrio de Hardy – Weinberg, en el cual se establece que en la primera generación de panmixia se alcanza el equilibrio, el cual se mantendrá en ausencia de selección, mutación, migración y deriva genética. Las frecuencias genotípicas en esta condición de equilibrio dependerán de las frecuencias génicas, y siempre existirán individuos heterocigotos si p ó q son distintos de cero (0).
| Generación de autogamia | % homocigosis | Genotipos | % heterocigosis |
| 0 | 0 | Aa | 100 |
| 1 | 50 | ¼ AA ½ Aa ¼ aa | 50 |
| 2 | 75 | 3/8 AA ¼ Aa 3/8 aa | 25 |
| 3 | 87,5 | 7/16 AA 1/8 Aa 7/16 aa | 12,5 |
| n | 100 | ½ AA ½ aa | 0 |
Cuadro 1. Efecto de la autogamia sobre los niveles de heterocigosis
El conocimiento del modo de reproducción debe ser una condición indispensable para iniciar un programa de mejoramiento genético, de tal manera de conocer la forma en que se pueda modificar la estructura genética de una población, que es el fin último de este tipo de programas.
Ing. Agr. Hernán E. Laurentin T. (M. Sc., Ph. D.)