



El concepto de fenotipo indica que la expresión de una característica en cualquier organismo vivo está relacionada con una proteína, que se formará al darse la transcripción de un gen particular para formar ARN mensajero, y el mensaje que él lleva como lenguaje nucleotídico será traducido a un lenguaje de aminoácidos para formar la proteína mencionada.
Para que se dé la transcripción de un gen, debe actuar sobre él una enzima llamada ARN polimerasa, la cual lee su información; y para actuar sobre él debe primeramente identificar al gen. Todo gen tiene un promotor, que es una secuencia de reconocimiento, de identificación del inicio del gen, secuencia que está presente en el ADN, físicamente unos cuantos nucleótidos antes que el gen inicie la secuencia nucleotídica que lleva el mensaje de la proteína a formar. El ADN está formado por pares de base (pb) que son numeradas en orden creciente desde el inicio de la región codificante; debido a que el promotor está antes del inicio del gen, sus pares de base (pb) son identificadas con un signo menos que las antecede.
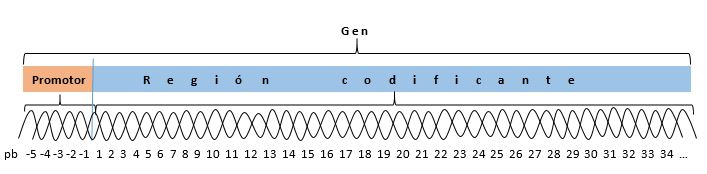
Para que se transcriba el gen 2, la ARN polimerasa debe leer dicho gen, y para eso primeramente debe identificar al gen y ubicarse sobre él. Para esto es el promotor, que tiene secuencias ubicadas a -35 y a -10 pares de base que le permiten a la ARN polimerasa identificar que allí existe un gen
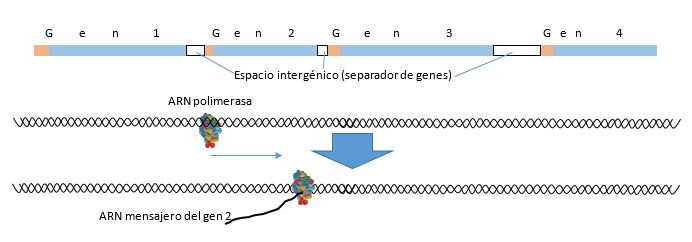
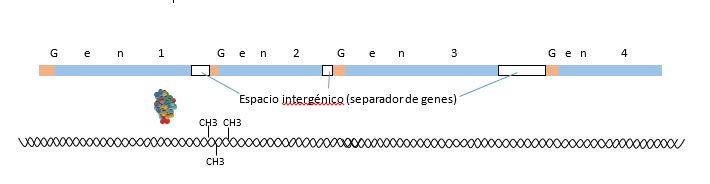
Muchos genes, quizás la mayoría o todos los genes, tienen los llamados mecanismos de regulación, que serán las distintas formas por la cual un gen dado no es “visto” por la ARN polimerasa, o lo contrario, es “permanentemente visto” por esta enzima. Si no es “visto” no se dará su transcripción y por tanto no habrá en la célula la presencia de la proteína que la transcripción y traducción de este gen originaría. Para no ser “visto” una de las estrategias conocidas hasta el momento es ubicar grupos metilo en la estructura de las bases nitrogenadas, especialmente sobre las citosinas del promotor del gen que se quiere ocultar. Cuando esto ocurre, la ARN polimerasa no reconoce a ese gen, por lo tanto no lo transcribirá, y por ende, no se logrará su traducción.
Para un mejor entendimiento de esto pudiéramos hacer el símil que el gen es una pista de aterrizaje y la ARN polimerasa es un avión; si se está dando un vuelo nocturno y la pista no está iluminada, el avión no logrará ver la pista y por tanto no aterrizará en dicha pista. Si el organismo o la célula “desea” que el gen sea “permanentemente visto” ubicará proteínas sobre su promotor para “avisar” a la ARN polimerasa que allí hay un gen para que sea transcrito. Estos mecanismos de regulación son conocidos desde hace años, y explican la razón por la cual un gen se transcribe y se traduce en un tejido dado, pero ese mismo gen no se transcribe ni se traduce en otro tejido. Por ejemplo los genes involucrados con la fotosíntesis se transcriben y se traducen primordialmente en tejidos foliares, pero no lo harán en tejidos de las raíces (esto partiendo del conocimiento de que en un organismo multicelular proveniente de la mitosis de una célula, bien sea por fusión de gametos o simplemente por reproducción asexual, posee exactamente la misma información genética, exactamente los mismos alelos, en todas las células que lo conforman).
Los mecanismos de regulación no se limitan a “ocultar” o “favorecer” el “avistamiento” de un gen actuando sobre el promotor, sino que este efecto también se logra actuando sobre las histonas, que son las proteínas sobre las que se ubica el ADN, y con las que, en conjunto con el ADN, forman los nucleosomas. Hasta hace relativamente poco tiempo se consideraba que las histonas eran simplemente soporte del ADN, pero ahora se conoce que no es un soporte pasivo, también pueden actuar como mecanismo de regulación de genes “ocultando” o “favoreciendo su avistamiento” por parte de la ARN polimerasa. En las histonas se logran estos efectos principalmente mediante la metilación y la acetilación.
Si la metilación se da en el promotor del gen, hay enzimas específicas que producen esa metilación. Esas enzimas específicas, como toda proteína en un organismo, provienen de la transcripción y la traducción de un gen, es decir, el ADN tiene en sí mismo genes con la información para regular la transcripción de otros genes, por lo tanto, hasta este momento no se hace referencia a la necesidad de utilizar un concepto distinto a la genética para explicar lo que sucede en el ADN. Sin embargo, la enzima que tiene la capacidad de metilar un promotor de un gen, lo puede hacer sobre cualquier gen, y dejar esa marca que oculte al gen a la vista de la ARN polimerasa sobre cualquier gen. Es aquí donde se da la necesidad de un concepto complementario a la genética, ya que es necesaria una explicación del por qué las marcas se dan sobre determinados genes, marcas que son altamente heredables, transmitidas de generación en generación. De forma similar, sea metilación o acetilación en las histonas, hay enzimas específicas que logran hacer esto sobre las histonas en cualquier sector de los nucleosomas, es por tanto necesario un nuevo marco conceptual que permita explicar por qué esto ocurre en ciertos y determinados sectores de ciertos y determinados nucleosomas, ocurrencia que es altamente heredable. Es a la dinámica de estas marcas que hace referencia el estudio de la epigenética, definida esta como los cambios heredables en la función del genoma sin cambios en la secuencia del ADN.
El entendimiento de la epigenética nos hace revisar el típico concepto de la genética en que la presencia de un alelo en un organismo, considerando las interacciones intra e interalélicas, es suficiente para la expresión de un fenotipo. Este concepto no es del todo cierto ya que conociendo lo que son las marcas sobre los promotores, ahora es necesario indicar que para que se exprese cierto fenotipo el alelo debe estar presente en el organismo y debe poder ser identificado dentro de la célula por la ARN polimerasa para que se dé su transcripción y posterior traducción.
La epigenética, a diferencia de la genética mendeliana, debe considerar en profundidad la influencia ambiental que se da sobre las marcas colocadas en los promotores o sobre las histonas. La genética mendeliana estudia caracteres definidos por solo un gen, por lo cual la influencia ambiental es prácticamente irrelevante, no así la epigenética. En el desarrollo de un organismo pluricelular la explicación del por qué un gen se expresa a partir de cierta edad del organismos, puede ser explicada por la epigenética. Genes que tienen que ver con la producción de pigmentos proteicos ubicados en los pétalos de la flor serán trascritos solo en las células que conforman los pétalos a partir de la madurez sexual del individuo, a pesar que ese gen está en todas las células del organismo durante toda la vida del mismo. Podría deducirse que sobre este gen, en todas las células, durante toda la vida del organismo, habrán marcas que impidan a la ARN polimerasa verlo, y por cambios en el ambiente celular de las células de los pétalos, estas marcas desaparecen en dichas células, permitiendo a la ARN polimerasa acceder a ese gen y transcribirlo. Como esos cambios no operan en el resto de las células, ese gen mantendrá las marcas que eviten a la ARN polimerasa identificarlo como un gen y transcribirlo. Si existen genes asociados a ciertas enfermedades o anomalías, no necesariamente el individuo que los porta sufrirá esa enfermedad, pues probablemente esos genes están marcados y nunca en la vida de ese individuo esos genes se transcribirán. Pero si el ambiente en que se desarrolla ese organismo es un ambiente que tiene la capacidad de alterar el ambiente celular de ciertas y determinadas células, pudiera darse el caso que las marcas desaparezcan permitiendo la transcripción del gen, su posterior traducción y por tanto la aparición de la proteína que permita el fenotipo de la enfermedad.
Ing. Agr. Hernán E. Laurentin T. (M. Sc., Ph. D.)