



La Agricultura constituye la mayor fuerza selectiva en la evolución de las malezas. Como consecuencia de haber desplazado la sucesión hacia estados tempranos en forma recurrente, las actividades agrícolas han mantenido las comunidades vegetales en estadios inmaduros. La mayoría de los componentes de esas comunidades son lo que en la agricultura llamamos malezas. De las 250.000 especies vegetales existentes, aproximadamente 8.000 (3%) son consideradas malezas y 250 sps. son problemáticas, representando el 0,1% de la flora mundial. El 70% de las malezas-problema corresponden a 12 familias botánicas y el 40% son pertenecientes a 2 familias: Poaceae y Asteraceae, presentándose la misma concentración de familias que en la situación de los cultivos más importantes.
Muchas de ellas se han introducido desde áreas geográficas muy distantes, o son nativas y particularmente favorecidas por las perturbaciones causadas en la actividad agrícola. Cualquiera sea su origen, las malezas son un componente integral de los agro ecosistemas y como tales influencian la organización y el funcionamiento de los mismos desde los albores de la agricultura.
Tradicionalmente, y principalmente debido a su impacto sobre el rendimiento, las malezas se han considerado organismos indeseables. Tanto en la literatura como en la tradición agrícola es muy profundo el sentimiento de aversión que estos organismos vegetales despiertan en el ser humano. En general, están asociadas a maldad, haraganería, daño, pérdida o inconvenientes de algún tipo.

Desde tiempos remotos y hasta épocas recientes, el problema de las malezas de los cultivos fue enfocado desde el punto de vista de su exclusión del cultivo. El esfuerzo por lograr ese objetivo ha sido descomunal y al tiempo que demuestra las habilidades del ser humano para desarrollar diferentes tácticas de eliminación o control, desnuda la ingenuidad con que ha sido enfocado el problema: salvo excepciones, las malezas de los cultivos son tanto problema en la actualidad como un siglo atrás.
El estudio de la ecofisiología de las malezas es, sin duda, la disciplina más joven de entre las componentes de la Protección Vegetal, si bien se ha desarrollado extraordinariamente en las últimas tres décadas. Es recién a partir de los años “70 que frente a la preocupación de la opinión pública por una mayor protección del medio ambiente y la aparición de algunos problemas derivados del uso abusivo de plaguicidas se forzó a todas las disciplinas encuadradas dentro de la P. Vegetal a tomar un enfoque integrado hacia el control de los diversos organismos infestantes. De esta forma se desarrollaron «Sistemas de Manejo Integrado» que, basándose en un conocimiento profundo de la biología y ecología de las malezas y de las posibilidades que ofrecen diversos métodos de lucha, permiten mejorar la eficacia en el manejo a la vez que minimizan el impacto ambiental.
La gran diferencia en la actualidad está basada en la gama de herramientas tecnológicas disponibles que permiten diseñar secuencias de cultivos con bajos niveles de infestación. Esto ha sido posible gracias a la investigación básica y aplicada relacionada con el estudio de los mecanismos fisiológicos de absorción, transporte y acción tóxica y al desarrollo, síntesis y producción de herbicidas. Se disponen hoy de más de 130 principios activos. El mercado mundial de agroquimicos moviliza una cifra superior a los 16000 millones de dólares por año: los herbicidas significan un 60 % de esta cifra y han contribuido en forma significativa al logro de los altos niveles de producción de las últimas décadas.
Hasta aquí, podemos concluir que los problemas de malezas de la actualidad son de similar envergadura que los existentes en el pasado y que la diferencia estriba en el rango de tecnologías que se disponen para enfrentarlas. Estas herramientas tecnológicas no son utilizadas en la mayoría de los casos con la eficiencia que demandan los tiempos actuales.
Una cuestión adicional se debe plantear: los grandes avances tecnológicos operados a partir de mediados del siglo XX han causado un cambio de importancia en las ideas centrales de la agricultura y han estado basadas en el uso intensivo de agroquimicos, de variedades e híbridos con altísimo potencial de rendimiento en condiciones de alta disponibilidad de recursos y en la reducción del requerimiento de trabajo humano a través del empleo generalizado de nuevas y potentes maquinarias y alto consumo de energía fósil. A este sistema se lo reconoce actualmente como agricultura convencional.
Durante los “70 varios autores han coincidido en expresar sobre las consecuencias de la concepción descripta y se plantearon posibles alternativas en el informe «Los limites del crecimiento», el cual si bien contenía algunas imprecisiones, planteaba la duda acerca de la sustentabilidad del sistema convencional.
La primera aproximación se ha ido enriqueciendo con información y evidencias que demuestran que con la tecnología promovida por la agricultura convencional ha ocurrido al menos en algunas áreas del planeta – una fuerte contaminación de aguas superficiales y subterráneas, se ha incrementado la erosión del recurso suelo, han aparecido formas de resistencia en plagas, y empiezan a registrarse residuos de plaguicidas en ciertos alimentos. Desde el punto de vista energético la agricultura convencional exhibe un balance de energía fuertemente negativo.
A partir de este diagnóstico, se han propuesto distintas alternativas tecnológicas que con diversas variaciones semánticas-sostenibilidad, sustentabilidad, agricultura orgánica, etc.- intentan el planteo más o menos amplio de nuevos sistemas de manejo que eliminen o al menos reduzcan la manifestación de los procesos antes enunciados.
Como regla general, las nuevas alternativas tecnológicas intentan la obtención de niveles de producción similares o algo inferiores a los actuales, sostenibles en el largo plazo, y con una sustancial reducción – en algunos casos del 50 %- en el uso de agro químicos, combustibles y labranzas.
Resulta claro que el nuevo enfoque no significa un retorno de los métodos anteriores a la Revolución Verde, sino básicamente en una combinación selectiva de las practicas proporcionadas por la tecnología moderna.
Si se tiene en cuenta que los ecosistemas bajo cultivo ocupan en la actualidad casi los dos tercios de la superficie terrestre y que contribuyen con el 20 % de la productividad neta del planeta, resulta claro que el protagonismo de los agricultores y técnicos que toman y ejecutan decisiones en tal sistema es de una envergadura superior a la de cualquier otro grupo social del planeta.
Por lo tanto, este replanteo de procesos productivos requiere necesariamente de productores y técnicos con un muy buen nivel de conocimientos acerca del sistema en que actúan. En síntesis, la toma de decisiones y la implementación de medidas y practicas que permitan la sustentabilidad de los sistemas agropecuarios es una cuestión de alta tecnología.
En ese contexto, el manejo de plagas animales y vegetales ocupa un lugar central.
Como se ha dicho en el comienzo, las malezas forman parte de un ecosistema altamente perturbado y en estado de etapa inicial de la sucesión secundaria perpetuo. El enmalezamiento posee además características diferenciales de la sucesión secundaria convencional:
- Existe un subsidio de energía al (fertilizantes, combustibles, agroquimicos).
- Hay recurrencia y cierta periodicidad en las perturbaciones (labranzas, herbivoría)
- El sistema produce información (modificaciones del ambiente térmico o lumínico) que es captada y almacenada por el banco de propágulos del
El banco de propágulos – semillas, frutos, rizomas, estolones, tubérculos y cualquier otra forma de propagación- constituye la pieza clave del proceso de regeneración de la vegetación. El estudio y la comprensión del funcionamiento de las poblaciones espontáneas que recurrentemente aparecen luego de la remoción del suelo o de la instalación de un cultivo constituyen un motivo central para determinar perfiles de decisión, dejando claro que las malezas no constituyen una clase botánica particular sino que son una población vegetal espontánea, que exhibe características propias para un sistema, en un determinado lugar y tiempo.
El tratamiento del problema de malezas en los cultivos bajo una óptica de corto plazo y en gran medida ignorando las características que exhiben la poblaciones espontáneas, ha demorado no solo la realización de estudios ecológicos aplicados al agro ecosistema sino la implementación de estrategias soportadas sobre la base de teoría ecológica disponible.
Resulta por lo tanto urgente no sólo la convergencia sino también la profundización de los estudios y la difusión de los conocimientos obtenidos en los agro ecosistemas a los fines de optimizar los esfuerzos realizados y compatibilizarlos con los desafíos de la agricultura sustentable.
- Las malezas: poblaciones exitosas en el agroecosistema
Las malezas son una forma especial de vegetación altamente exitosa en ambientes agrícolas: son poblaciones vegetales que crecen en ambientes perturbados por el hombre sin haber sido sembradas.
Desde un punto de vista ecológico las malezas pueden ubicarse dentro de las pioneras de la sucesión secundaria. En el agro ecosistema el impacto más crítico de las malezas es el efecto negativo sobre las plantas cultivadas ejercido a través de la competencia por recursos limitados y las alelopatías. Trastornos en la recolección y el acondicionamiento de los granos y la disminución de la calidad del forraje constituyen perjuicios adicionales en muchos sistemas.
Éxito, desde el punto de vista evolutivo, implica generalmente la perdurabilidad o continuación de la linea genética a través del tiempo. De esta forma el éxito evolutivo esta reflejado por el número de individuos, la Capacidad Reproductiva, el área y el rango de hábitats ocupados por la especie en cuestión.
La comprensión acerca del concepto de éxito en el agroecosistema es necesario para explorar la naturaleza del enmalezamiento y sentar bases sólidas de manejo en los sistemas de producción.
- Definición de Éxito.
Las malezas más exitosas en ecosistemas agrícolas son a menudo las que se consideran más problemáticas. El éxito puede medirse en este contexto, según la rapidez de colonización, la dificultad de su eliminación y el efecto negativo sobre la productividad de las especies cultivadas.
En general, la gran mayoría de las características de una planta que contribuyen a una alta capacidad de colonización y competitividad se corresponden con las exhibidas por las malezas, y en un grado mucho mayor que el exhibido por las especies cultivadas.
- Estrategias adaptativas de las malezas
Una aproximación comúnmente utilizada por los ecólogos con el fin de comprender las relaciones entre los organismos y su ambiente es la de comparar sus estrategias adaptativas. Las poblaciones se adaptan al ambiente mediante el balance de asignación y partición de recursos y energía entre los procesos fisiológicos y componentes de la biomasa a lo largo del ciclo de vida.
En otras palabras, debe existir una solución de compromiso entre los recursos que se asignan para competir exitosamente en el presente (biomasa asignada a estructuras vegetativas) y aquellos que se destinan al futuro, es decir a la descendencia (biomasa asignada a semillas). De este concepto surge la idea de “Valor Reproductivo” una cuestión central en una población exitosa: el conjunto de características de la historia de vida que maximizan la supervivencia y reproducción configuran la adecuación, en un ambiente particular. Las poblaciones de malezas exhiben una adecuación elevada o máxima.
- La selección r-K.
Una teoría ampliamente aceptada y que concierne con las estrategias de especialización evolutiva es la de la selección r-K. Mientras que los organismos r seleccionados están adaptados para la colonización y la reproducción en ambientes impredecibles, los organismos K seleccionados están adaptados para la persistencia y la reproducción en ambientes estables. Las poblaciones r seleccionadas ocupan los estadios iniciales de la sucesión, en cambio las poblaciones K seleccionadas ocupan mayormente los estadios más tardíos de la sucesión.
Si bien muchas de las poblaciones de malezas exhiben características propias de la estrategia r, otras poblaciones pueden ubicarse dentro de K y en muchos casos las poblaciones exhiben características comunes a ambas ubicándose en un continuum entre r y K.
Altieri et al.,1988 sugieren una hipótesis alternativa en relación con las estrategias adaptativas y que definen más claramente el rol del ambiente en determinar el éxito de la planta. Proponen que los factores externos que limitan la cantidad de biomasa que puede generarse en un ambiente dado pueden ser clasificados en dos categorías: el primero es el estrés y el segundo es el disturbio.
Si bien estos dos factores varían entre los hábitats, los casos extremos pueden combinarse de tal manera que resultan en tres tipos de hábitats y tres combinaciones posibles de estrategias evolutivas.
Las Competitivas maximizan la captura de recursos en condiciones altamente productivas y no perturbadas. Son abundantes durante las etapas tempranas e intermedias de la sucesión. Las Tolerantes a estrés son plantas longevas adaptadas para vivir en condiciones de productividad limitada y a menudo ocupan los estadios tardíos de la sucesión. Las Ruderales, son a menudo plantas de vida muy corta con alta producción de semillas y que ocupan los estadios muy tempranos de la sucesión.
- Habilidad competitiva
Las secciones precedentes han demostrado que las malezas están adaptadas al ambiente agrícola a través de una serie de estrategias que maximizan un rápido crecimiento y una reproducción prolífica en hábitats perturbados. Estas características también contribuyen a su competitividad.
La habilidad competitiva de las malezas es una función compleja donde se combinan características que resultan en un rápido agotamiento de los recursos necesarios para el cultivo.
Ciertas características están recurrentemente asociadas con la habilidad competitiva: entre ellas se incluye a una gran cantidad de reservas acumuladas en órganos de propagación vegetativa o almacenaje que conduce a una rápida expansión del follaje, un sistema aéreo y subterráneo vigoroso y de rápido crecimiento que permite un rápido aprovechamiento de los recursos del ambiente y una expansión tanto lateral como horizontal que resulta en una muy alta densidad de vástagos y raíces. La fenología es otra característica de la habilidad competitiva.
- Características de las malezas asociadas al éxito ecológico.
- Clasificación y características de las Malezas Cosmopolitas.
Si consideramos una cifra superior a 250.000 especies vegetales dentro de las Fanerógamas, menos de 250 se consideran importantes en amplios sectores del planeta. De estas unas 70 se consideran «:las peores del mundo». Estas 70 especies están distribuidas en 30 familias, pero casi los dos tercios están ubicadas en 8 familias y alrededor del 50 % están ubicadas en solo dos familias :Poáceas y Asteráceas. El bajo número de especies consideradas maleza en relación al total, ejemplifica la fenomenal presión selectiva ocurrida desde tiempos remotos.
De las 30 familias botánicas que contienen a las peores malezas del mundo, 5 de ellas -Poáceas Solanáceas, Convolvuláceas, Euforbiáceas y Fabáceas también suministran el 75 % del alimento mundial. Esta observación implica que los cultivos y las malezas comparten características taxonómicas y orígenes evolutivos comunes.
Se describen a continuación características eco fisiológicas que exhiben las plantas que los seres humanos designamos como malezas.
- Características de la Fase Regenerativa.
Los eventos que suceden en la fase regenerativa de las plantas (dispersión de las semillas, dormición, germinación y establecimiento de las plántula) son particularmente importantes para las malezas en ambientes agrícolas.
- Dispersión de las semillas en el espacio
La dispersión es la diseminación de los propágulos de una planta desde un lugar a otro. La significancia para un organismo de este proceso depende de su ambiente. Tanto para las malezas como para otras especies que conforman la sucesión, la supervivencia continuada depende del escape y el establecimiento a nuevas áreas que puedan ser más benignas para la supervivencia que la ocupada por su progenitor.
Además del número de semillas que cada individuo produce, otros factores importantes que determinan el número de semillas que caen por unidad de área son la dispersabilidad de las semillas y la actividad de los agentes de dispersión.
La dispersabilidad es una función de las características de las semillas, las cuales incluyen apéndices sobre la semilla o el fruto. Muchas malezas comunes poseen formas muy especializadas de dispersión mecánica, mientras que en otras la semilla es propulsada por diversos mecanismos de tensión que ocurren en el fruto. La forma más común de dispersión es la pasiva, la cual se incrementa por la actividad de los agentes de dispersión como el viento, agua, animales y el hombre.
Si bien en algunos casos los agentes de dispersión naturales constituyen el mecanismo principal (caso del viento y las semillas provistas de pappus en Carduus sp, muy livianas como en el caso de la parásita Orobanche sp o transportadas por la planta entera como en Salsola kali) las actividades del hombre constituyen el aporte más significativo en este aspecto. La siembra de semilla sin certificación de pureza, la falta de limpieza en los herramientas e implementos de labranza y cosecha, el traslado de hacienda y el agua de riego constituyen entre otras las principales vías de dispersión e invasión de malezas en nuevas áreas. Las normas, regulaciones y leyes de orden sanitario implementadas por los gobiernos de todo el mundo apuntan a disminuír este efecto. En el cuadro 1 se reseña la alta y variable producción de semilla de algunas malezas, así como su viabilidad en el suelo.
Cuadro 1- Producción y viabilidad de semillas de algunas malezas importantes en la agricultura
|
Maleza |
Producción de semillalplanta |
Viabilidad en el suelo (años) |
|
Avena fatua
Avena guacha |
100-450 |
3-8 |
|
Senecio vulgaris
Senecio |
1100-1200 |
|
|
Capsella bursa-pastoris
Bolsa de pastor |
3500-4000 |
16-35 |
|
Portulaca oleracea
Verdolaga |
10000 |
30-40 |
|
Stellaria media
Capisquí |
15000 |
|
|
Echinochloa crus-galli
Capín |
2000-150000 |
|
|
Solanum nigrum
Tomatillo |
178000 |
>39 |
|
Chenopodium album
Chenopodio |
13000-500000 |
>39 |
Adaptado: Cobb,A.,1992.
- Dormición y Germinación de las Semillas
La mayoría de las malezas exitosas poseen prolongada viabilidad y pronunciada dormición, permitiendo su supervivencia en condiciones inadecuadas para el crecimiento de las plantas y la persistencia por largos períodos en el suelo. La dormición puede ser definida como un estado en el cual las semillas o yemas viables no germinan aún cuando existan condiciones de temperatura, humedad y concentración de oxígeno adecuadas.
Se denomina germinación al proceso por el cual el embrión reasume el crecimiento, lo cual resulta en su emergencia a través de las cubiertas de la semilla o fruto. La dormición está considerada como el factor primario que contribuye a la presencia continuada de semillas de malezas en los suelos agrícolas. Como contraste, la dormición ha sido eliminada por mejoramiento genético en la mayoría de las especies cultivadas de tal manera que ocurra una germinación y emergencia sincrónica.
Que una semilla se duerma o germine depende de una serie de factores relacionados con la característica de la semilla y factores ambientales. Estos dos componentes interactúan de tal manera que la dormición y la germinación son procesos fuertemente sincronizados con los cambios ambientales. La sincronización ambiental provee a las semillas de una mayor seguridad de supervivencia, dado que la germinación ocurre en el momento en que las condiciones ambientales son también favorables para el crecimiento.
La dormición es una característica muy variable y aún entre las semillas de la misma planta. Este polimorfismo somático asegura la supervivencia continuada de las especies por reducción del riesgo de pérdida total en el caso que las condiciones luego de la germinación sean desfavorables para el establecimiento. El polimorfismo es muy común en las familias Asteráceas, Brasicáceas, Chenopodiaceas y Poáceas.
Si bien la dormición ha sido estudiada en detalle en numerosas especies, la fisiología de este proceso no está totalmente comprendida. Generalizadamente, se puede decir que la dormición y la germinación son procesos muy complejos e interrelacionados, que están controlados por factores bioquímicos, fisiológicos y estructurales.
El rol del ambiente en la regulación de la dormición y su desbloqueo – esto es la germinación -se extiende hasta el período de maduración cuando la semilla está en desarrollo en la planta progenitora. Las semillas que están dormidas cuando se desprenden de la planta se designan con dormición innata o primaria. La dormición innata puede estar causada por una inmadurez o exceso de humedad en el embrión o un requerimiento de postmaduración conferido por un régimen especial de temperatura, luz o estratificación.
Las semillas que no germinan debido a la restricción impuesta por un factor ambiental se designan como quiescentes o con dormición impuesta.
Las semillas no dormidas y que entran en dormición luego de la dispersión a menudo en respuesta a condiciones ambientales que inhiben la germinación se consideran con dormición inducida o secundaria.
Las semillas presentes en el perfil del suelo presentan ciclos de dormición/no dormición que promueven la germinación durante el período más favorable para el desarrollo de la plántula. Esta compleja estrategia puede ser ejemplificada con el comportamiento de Ambrosia artemisiifolia, una especie propia de los estadios sucesionales tempranos.
Cuando las semillas caen al suelo se encuentran dormidas y germinan luego de la estratificación invernal siempre y cuando el suelo sea perturbado y las semillas sean llevadas a la cercanía de la superficie del suelo. Las semillas que no germinan, pueden inducirse a una dormición secundaria. Esta dormición puede ser desactivada por otro ciclo de estratificación.
La germinación de las malezas y otras especies de la sucesión temprana a menudo requieren luz para germinar pero son inhibidas por la luz filtrada por la vegetación (rojo lejano) y promovidas por temperaturas fluctuantes. Estas condiciones se cumplen cerca de la superficie de suelo desnudo donde los niveles de irradiancia son altos y las amplitudes térmicas elevadas.
La germinación de semillas de malezas también se ve muy disminuida con la profundidad, donde los niveles de oxígeno son bajos, hay ausencia de luz y temperaturas menores.
La mayoría de las semillas de malezas germinan mejor a profundidades de 0.5 a 2 cm. La adaptación a germinación cerca de la superficie asegura la disponibilidad de recursos y reduce la probabilidad de competencia con las especies de germinación y establecimiento más tardío. De esta manera, la germinación de semillas de malezas es promovida por las labranzas, que traen semillas ubicadas en profundidad hacia la superficie.
Una práctica común de manejo es la de traer semillas del estrato profundo al superficial mediante labranzas, esperar la ocurrencia de lluvias antes de la siembra removiendo las plántulas de malezas con nuevas labranzas o herbicidas. Al revés, en sistemas con mínima labranza, la ausencia de perturbación del perfil del suelo asegura que las semillas permanezcan en los estratos profundos, donde no se generan condiciones aptas para germinación, perdiendo en el transcurso del tiempo viabilidad y en este sistema las malezas perennes irán ocupando lugares de mayor importancia.
- Germinación y establecimiento de plántulas.
Cuando la dormición concluye, la secuencia normal de eventos conduce a la emergencia de la radícula, el crecimiento subterráneo de la raíz primaria, la emergencia del vástago por encima del suelo y finalmente al establecimiento de la plántula. El establecimiento concluye cuando la plántula es capaz de realizar un crecimiento independiente de las reservas de los cotiledones.
La emergencia en la mayoría de las dicotiledóneas es epígeo, es decir que los cotiledones se elevan por encima de la superficie del suelo. En la emergencia hipogea los cotiledones permanecen debajo de la superficie del suelo. Luego de la emergencia, los cotiledones adquieren rápidamente su condición de órganos fotosintetizantes y el área foliar se agranda rápidamente.
Varios autores proponen el concepto de «Sitio seguro» y lo definen como la zona en que se encuentran condiciones apropiadas para la germinación y la supervivencia. El sitio seguro provee entonces no sólo estímulos para desbloquear la dormición y favorecer la germinación sino también ausencia de peligros como predatores, competidores, patógenos y toxinas.
A los propósitos del manejo de malezas, resulta imperativo distinguir a las plántulas – provenientes de semillas- de los brotes o expresiones vegetativas provenientes de órganos de multiplicación vegetativa como rizomas, estolones o tubérculos. La plántula representa el estadio más vulnerable en el ciclo de vida de la planta y generalmente es el más fácil de ser destruído ya sea mecánicamente o con herbicidas.
- Características de la Fase Establecida
La fase establecida de las plantas involucra numerosas funciones relacionadas con la captura de recursos, crecimiento y reproducción. Los eventos de esta fase son los mayores determinantes del éxito competitivo y de la supervivencia en el agro ecosistema. En términos de reproducción, las malezas tienen sistemas de intercambio genético variables y generalistas, producen abundante cantidad de semillas, – por un largo periodo de tiempo y se reproducen vigorosamente a partir de fragmentos si son perennes. Estas características contribuyen en forma significativa al éxito en ambientes perturbados.
- Orden de emergencia y Capital inicial
El éxito de una planta esta asociado con la rapidez con que se inician los procesos de germinación y la velocidad con que se genera biomasa adicional.
Está demostrado que las semillas que germinan primero y que poseen mayor capital inicial (embriones mayores o tejidos de acumulación y reserva más abundantes) poseen una clara ventaja sobre los individuos de germinación más tardía o con menor capital inicial. Como se explica más adelante, la ocupación del espacio y la captura de recursos limitantes constituyen la clave de una población exitosa en la fase de establecimiento. Resulta ilustrativo observar el cuadro 2 en donde se plantean las tasas máximas de crecimiento relativo semanal de diferentes especies de malezas problemáticas en condiciones de competencia con cultivos.
Cuadro 2.- Tasas de crecimiento relativo potenciales, de diferentes malezas presentes en cultivos y praderas.
|
Especie |
Nombre común |
Tasa de crecimiento R máx (g/g.semana) |
|
Chenopodium albun L. |
Chenopodio |
2,12 |
|
Convolvulus arvensis L. |
Corrihuela |
2,44 |
|
Dactylis glomerata L. |
Pasto miel |
1,31 |
|
Festuca rubra L. |
Festuca |
1,18 |
|
Matricaria matricarioides |
Manzanilla |
1,17 |
|
Plantago lanceolata |
Llantén |
1,70 |
|
Poa annua L. |
Pastito de invierno |
2,70 |
|
Polygonum aviculare |
Pasto alambre |
1,43 |
|
Senecio vulgaris L. |
Senecio |
1,63 |
|
Stellaria media |
Capiquí |
2,43 |
|
Yaraxacum officinale |
Diente de león |
1,19 |
Adaptado: Radosevich, S.R. et al.; 1984.
- Crecimiento y Captura de de Recursos
En general, los individuos que crecen más rápido que sus vecinos utilizan una fracción desproporcionada de los recursos disponibles, en detrimento de aquellos. La Tasa de Crecimiento Relativo (RGR) ha sido usada como medio de integrar las características fisiológicas en un valor común que permita comparar diferentes especies.
Las plantas anuales tienen con frecuencia valores de RGR mayores en relación con plantas leñosas. En términos de estrategia adaptativa, tanto las Competitivas como las Ruderales poseen RGR elevadas. Esta característica está altamente correlacionada con una explotación oportunista de los hábitats perturbados y también constituye una tendencia general entre las especies que conforman los estadios sucesionales tempranos.
Un componente importante del crecimiento de la planta es la partición de fotosintatos hacia la generación de área foliar adicional. Se demostró que la partición del área foliar – o la tasa de expansión de área foliar nueva – estaba altamente correlacionada con un rápido crecimiento, Radosevich, et al, 1984; demostraron que los parámetros de crecimiento relacionados con el tamaño de la plántula y el área foliar fueron los mejores predictores de la competitividad en mezclas de malezas. Resulta claro que la rápida producción de área foliar, que se traduce en un rápido desarrollo del canopeo, es crítica para el éxito tanto de malezas como de cultivos en el ambiente agrícola.
Así como en la fase regenerativa, los eventos de la fase establecida están muy ligados a los factores ambientales. Los recursos, incluyendo la luz, agua, nutrientes minerales, CO2 y O2 son factores consumibles del ambiente que pueden limitar el crecimiento cuando están ausentes o en déficit. Como contraste, si bien las condiciones ambientales como temperatura, pH y densidad aparente del suelo no son consumidos, tienen una crucial influencia sobre los procesos que determinan el crecimiento de la plántula. La capacidad diferencial de las plantas de extraer recursos o de producir respuestas distintas a las condiciones ambientales son los principales determinantes del éxito competitivo entre especies.
Los procesos de particular importancia para el éxito de las malezas en ambientes agrícolas incluyen el uso eficiente del CO2, la rápida y eficiente absorción de agua a través del sistema radical, la alta eficiencia en el uso del agua y la plasticidad morfológica y fisiológica en relación con cambios ambientales.
- Ciclos Fotosintéticos.
Basadas en sus ciclos fotosintéticos, las plantas pueden dividirse en tres grupos principales. Estos incluyen C3 (Ciclo de Calvin-Benson), C4(Ciclo Hatch- Slack) y CAM.
Si bien en cada uno de estos tres grupos se pueden encontrar a varias malezas, la vía C4 es la más representada por las malezas principales. Otros estudios han demostrado que sólo el 0.4 % de la flora posee esta vía adicional de incorporación de CO2. De las 76 malezas más importantes del mundo 42 % emplean la vía C4 y 78 % de las 18 malezas más agresivas son C4. Un estudio similar ha demostrado que 16 especies cultivadas tienen la vía C3.
Las plantas que poseen la vía C4 generalmente tienen tasas de fotosíntesis neta más elevadas que las plantas C3. La enzima que inicia la fijación de CO2 [PEP (fosfonenolpiruvato)carboxilasa] tiene una afinidad más alta por el CO2 que el de la RuBP (ribulosadifosfato) carboxilasa, enzima que inicia la fijación de CO2 en el Ciclo C3. En la vía C4 el CO2 es concentrado en las células del mesófilo y pueden mantenerse altas tasas de fotosíntesis, aún cuando los estomas están prácticamente cerrados o cuando la concentración de CO2 en el aire es baja. Además, dada la alta concentración de CO2 interna, la fotorespiración disminuye sustancialmente, lo cual contribuye para el logro de altas tasas de fotosíntesis neta. Debe puntualizarse finalmente que en las plantas C4 funcionan en realidad ambos ciclos acoplados, actuando el ciclo Hatch-Slack como reservorio de CO2 fijado.
La alta eficiencia de fotosíntesis está a menudo correlacionada con un rápido crecimiento, el que puede conferir una habilidad competitiva superior. Esto llevó a varios autores a clasificar a las plantas según un criterio de eficiencia o no eficiencia basado en sus características fotosintéticas (C3=ineficientes, C4=eficientes). Investigaciones posteriores han restado generalidad a esta idea, dado que especies C3 son altamente competitivas. Es que otros factores concurren para determinar la habilidad competitiva de una planta. Por ejemplo, para la mayoría de las plantas C4 la temperatura óptima para la fotosíntesis y el crecimiento es más alta que para las plantas C3. En un hábitat de condiciones térmicas más templadas la ventaja de las especies C4 no es tal, no sólo porque la tasa fotosintética total es menor sino también porque la vía acoplada C3-C4 tiene un mayor requerimiento energético para funcionar. En ambientes agrícolas caracterizados por estrés hídrico y elevadas temperaturas muy probablemente la mayoría de las malezas anuales y perennes de verano van a pertenecer al grupo C4, mientras que las anuales de invierno serán a menudo especies C3; situación constatada en nuestras condiciones de producción.
- Absorción de Agua
Además del suministro estacional del agua, las plantas pueden regular la disponibilidad de agua en cualquier ambiente mediante una serie de mecanismos y procesos. Estos son: el desarrollo, estructura y distribución de raíces, tolerancia a bajo potencial de agua en los tejidos, control de pérdidas de agua por transpiración, y eficiencia en el uso del agua.
La estructura de las raíces 20 días post-emergencia de la mayoría de las malezas tienen sistemas radicales mayores y superficies de absorción más elevadas que los cultivos con los cuales compiten. El desarrollo del sistema radical es el principal factor que contribuye a una explotación de la humedad del suelo y los nutrientes asociados, confiriendo así una competitividad superior.
El control fisiológico de la disponibilidad de agua por la planta puede ser expresado como la Eficiencia en el uso del Agua, que se expresa por la cantidad de CO2 fijado o la cantidad de materia seca producida, por unidad de agua perdida por transpiración, en el cuadro 3 se observa las necesidades de agua comparativamente entre algunas malezas y cultivos, necesarios para la formación de materia seca, las plantas difieren significativamente en sus requerimientos de agua, constatándose que los valores más altos de la eficiencia en el uso de agua (cifras menores en el cuadro 3) estaban en especies C4 mientras que las especies C3 tenían valores relativamente más bajos. De esta manera, las plantas C4 son más productivas que las C3 cuando el agua es limitante.
Cuadro 3.- Necesidades de agua para la formación de materia seca y vía fotosintética de diferentes malezas y cultivos
|
Especie |
Nombre común |
Vía fotosintética |
g H2Olg M.S. |
|
Zea mays |
Maíz |
C4 |
250 |
|
Portulaca oleracea |
Verdolaga |
C4 |
281 |
|
Amaranthus spp. |
Yuyo colorado |
C3 |
290 |
|
Sorghum |
Sorgo |
C4 |
304 |
|
Xanthium sp |
Abrojo |
C3 |
415 |
|
Chenopodium album |
Chenopodio |
C3 |
435 |
|
Triticum aestivum |
Trigo |
C3 |
542 |
|
Solanum tuberosum |
Papa |
C3 |
575 |
|
Citrullus vulgaris |
Sandía |
C3 |
577 |
|
Avena sativa |
Avena |
C3 |
583 |
|
Polygonum aviculare |
Pasto alambre |
C3 |
678 |
|
Oryza sativa |
Arroz |
C3 |
682 |
|
Cucumis sativus |
Melón |
C3 |
683 |
|
Phaseolus vulgaris |
Poroto |
C3 |
700 |
Adaptado: Black, et al.;1969 y Cobb,A.;1992.
El rápido desarrollo radical asociado con un uso excesivo del agua contribuyen en forma significativa tanto al enmalezamiento como a la competitividad. La ventaja conferida por la vía fotosintética C4 y los valores asociados de alta eficiencia del uso de agua que se han encontrado en muchas malezas serían más importantes en ambientes más cálidos y secos y con elevada irradiancia y donde ocurren con gran frecuencia situaciones de estrés hídrico.
Una cuestión adicional se demuestra en el caso de Xanthium sp (abrojo), una maleza muy frecuente en diferentes cultivos, se ha comprobado que tanto en los estadios vegetativos como reproductivos de los cultivos, el nivel de potencial agua del xilema es siempre más negativo en Xanthium. Paralelamente, el proceso de mejoramiento genético de los cultivos, que ha venido jerarquizando la mayor producción de la porción comercial parece haber descuidado este aspecto.
- Plasticidad y Respuestas ambientales
Los ambientes agrícolas en los cuales se encuentran las malezas son a menudo sistemas muy perturbados en los cuales se presenta el suelo altamente expuesto, sin cobertura con extremos de temperatura en su superficie y fluctuaciones importantes tanto en la humedad como en los niveles de nutrientes. Estos cambios generalmente ocurren de manera impredecible.
Las malezas mejor adaptadas a estas condiciones son tolerantes a estas condiciones variables de tal manera que pueden crecer y reproducirse en forma exitosa. A esta tolerancia a la variación ambiental se la designa plasticidad.
La plasticidad es la capacidad de un ajuste fenotípico -traducido en variaciones morfo y fisiológicas – condiciones cambiantes. Las malezas poseen un genotipo de todo uso o multipropósito, lo que en otras palabras implica la existencia de un amplio rango de caracteres exhibidos por un limitado número de genotipos. Existen ideas distintas sobre este punto. Más bien se puede considerar a las malezas como poblaciones que contienen genotipos que responden a un alto grado de especialización y con un grado de ajuste muy fino a la oferta de señales que realiza el ecosistema bajo cultivo.
Lo que sí es incuestionable es que las especies cultivadas han estado sujetas a un intensivo proceso de mejoramiento y a través de él han perdido flexibilidad morfológica y fisiológica para responder a variaciones ambientales.
Un ejemplo sobresaliente de la plasticidad de las plantas es la variación del número de sus partes o módulos. Otras variaciones plásticas incluyen el tiempo que transcurre entre diversas etapas del ciclo de crecimiento (emergencia- floración o floración-maduración), el tipo de semillas que producen (nivel de dormición) y el tamaño relativo y distribución de raíces y hojas.
Las variaciones en el número, el tamaño y la distribución de las partes aérea y subterráneas de la planta reflejan respuestas plásticas de la asignación y partición de fotosintatos. Todos estos procesos confieren a una maleza la capacidad de rápida respuesta ante cambios bruscos en la distribución de recursos del hábitat.
Las malezas también exhiben una respuesta plástica a la densidad. Una densidad creciente impone un estrés entre las plantas como consecuencia de la disminución de los recursos ambientales, una cuestión que se magnifica en la medida que las plantas crecen. A medida que las plantas crecen, la mortalidad densodependiente y el consiguiente aumento de tamaño de los sobrevivientes – crecimiento plástico- conduce a una situación en donde sobreviven pocos individuos pero muy grandes. En términos generales se puede afirmar que este es un proceso de amplia ocurrencia y esta designado como Ley del Auto raleo, citada por varios autores.
Como consecuencia de los procesos mencionados, se presentan situaciones donde existen muchos individuos pero pequeños y otras donde existen pocos pero más grandes. La expresión de la biomasa vegetal por unidad de área conduce a otra generalización: en un amplio rango de densidades, la biomasa por unidad de área es constante.(Ley del Rendimiento Final Constante) y decidida por factores de mayor jerarquía como la latitud, irradiancia, disponibilidad de agua y nutrientes en el suelo.
- Floración y Producción de semillas
El sistema de mejoramiento de las malezas constituye uno de los componentes cruciales del éxito. La mayoría de las malezas exhiben un alto nivel de endocría ya sea a través de la autogamia (autopolinización) o mediante la agamospermia. No obstante en todas ellas existe un nivel de alogamia, lo cual permite el intercambio de material genético con otras poblaciones. La alogamia es producida generalmente por el viento o visitantes generalistas de la flor más que por polinizadores específicos.
Este sistema combinado conduce a la producción de duplicados estables del genotipo padre, pero surgidos de recombinación y variabilidad genética amplia.
Las ventajas de este sistema combinado son evidentes: el testeo de una amplia gama de genotipos a los efectos de que uno de ellos logre el éxito y el rápido avance posterior del genotipo más adecuado. Este esquema resulta aun mas eficiente en las especies que poseen propagación vegetativa.(Sorghum halepense, por ejemplo).
Las malezas están caracterizadas por una producción de semilla abundante y continuada bajo condiciones ambientales diferentes. También la densidad juega un papel crucial en la producción de semillas por cada individuo: la fecundidad es un atributo fuertemente densodependiente.
Dos malezas muy comunes en cultivos de primavera verano, como Amaranthus quitensis y Datura ferox muestran claramente esta respuesta. En este último caso, esta característica poblacional, asociada a prácticas de manejo de cultivos provocan una redispersión de semillas de la maleza en el campo, permitiendo explicar la presencia de chamico aún cuando la eficiencia de los herbicidas utilizados para su control es muy elevada.
- Características Especiales de las Malezas Perennes
La mayoría de los atributos de las malezas previamente descriptas se aplican a las malezas perennes. Esto es especialmente cierto en el caso de especies perennes muy prolíficas como Cyperus rotundus. La reproducción vegetativa vigorosa y la regeneración a partir de fragmentos constituyen otra de las características esenciales en las malezas perennes.
La significancia de la propagación vegetativa en la lista original es importante: el 61 % de las 18 peores malezas exhibe alguna forma de propagación vegetativa. Como se ha explicado antes, la propagación vegetativa tiene los mismos efectos que la autopolinizacion o retrocruza: el resultado de ambos procesos es una rápida multiplicación de individuos con genotipos apropiados para el ambiente en cuestión. El balance entre la propagación vegetativa y la reproducción por semillas en cualquier especie es altamente dependiente de las condiciones ambientales, incluyendo la densidad.
En el caso de las especies que poseen ambos medios de multiplicación – sexual y asexual – como en Sorghum halepense, la combinación selectiva de los dos procesos exhibe una complementariedad muy eficiente: la reproducción sexual es significativa en la etapa de colonización o de reinstalación de la plaga en el campo y la propagación vegetativa multiplica muy rápidamente los genotipos más exitosos.
Las plantas que se reproducen vegetativamente poseen estructuras especializadas que pueden sobrevivir en temperaturas bajas, algún nivel de sequía u otra condición desfavorable. Cuando estas estructuras están localizadas por debajo de la superficie del suelo son muy persistentes y robustas. La función y significancia de las estructuras reproductivas vegetativas es doble: por un lado producen yemas o meristemas que pueden generar vástagos y raíces y por el otro sirven como tejidos de almacenaje y reserva de carbohidratos.
A menudo, más de un tipo se encuentra en la misma planta. Estas estructuras incluyen a estolones, rizomas, tubérculos, turiones, bulbos, cormos, coronas y raíces gemíferas. Cyperus rotundus, por ejemplo, posee rizomas, bulbos y tubérculos. De estos, el último contribuye significativamente a la propagación de la maleza.
La habilidad competitiva de una maleza perenne está significativamente aumentada por la cantidad de carbohidratos de reserva almacenados en las estructuras subterráneas. La rápida expansión del canopeo, factor principal que contribuye a la habilidad competitiva de muchas malezas perennes, ocurre debido a la movilización de las reservas de energía y materiales estructurales acumulados en los órganos de reserva originados en la estación de crecimiento anterior. Estas reservas preexistentes permiten una rápida emergencia y crecimiento de los jóvenes vástagos que poseen así una ventaja competitiva sobre aquellos que provienen de semillas, en general más pequeñas y con menor capital inicial.
La cantidad de carbohidratos almacenados en los órganos subterráneos varían en relación con la época del año. En las épocas más desfavorables se produce una disminución debido al consumo por respiración de mantenimiento y en la etapas iniciales de la brotación como consecuencia del abastecimiento a los jóvenes vástagos en crecimiento. En las épocas favorables, el sistema aéreo produce fotosintatos que son trasladados a nuevas estructuras subterráneas.
El estudio de las variaciones en el nivel de biomasa aérea y subterránea y de carbohidratos resulta de sumo interés no sólo para comprender los procesos de interacción maleza-cultivo sino también para aumentar la eficiencia de las tácticas de control. La búsqueda de predictores de fácil medición para estas variaciones constituyen lineas de trabajo de suma importancia práctica,por ejemplo determinar localmente el momento de inicio de tuberización en el caso de Cyperus rotundus.
La propagación vegetativa puede estar acompañada por una fragmentación de órganos tanto aéreos como subterráneos (ej.; estolones y rizomas de Cynodon dactylon). La división de las estructuras vegetativas puede ocurrir espontáneamente debido a la muerte de los tejidos de conexión, a la muerte de la planta madre o a una labranza: sin ninguna duda este último evento es el más eficiente para dispersar los propágulos de la maleza. Luego de la separación, las plantas que resultan de cada propágulo vegetativo son clones de la planta madre lográndose así una propagación horizontal del genotipo.
En algunos casos, las propágulos vegetativos exhiben dormición o se manifiestan fenómenos de correlación inhibitoria (dominancia apical), pero en la gran mayoría la ausencia de crecimiento de las yemas o meristemas esta asociada a restricciones ambientales (sequía o temperatura). En el caso de la temperatura, existe un umbral de brotación variable para especies y aún ecotipos.
Cualquiera sea el mecanismo específico, las estructuras regenerativas de las plantas que almacenan reservas y que tienen la capacidad de propagarse vegetativamente son componentes significativos en los problemas de malezas de los agroecosistemas.
- Competencia entre cultivos y malezas
La competencia juega un rol central en el balance productivo de los agroecosistemas, la estructura y productividad de la comunidad se halla fuertemente influenciada por las relaciones competitivas entre los distintos componentes. La competencia puede definirse como una interacción entre individuos, provocada por la demanda común de un recurso limitado, y que conduce a la reducción de la performance final de esos individuos.
Los recursos involucrados clasicamente en condiciones de campo, cultivo y malezas van a competir por luz, agua y nutrientes. A pesar de la importancia teórica y práctica de definir en cada situación cual es él o los factores involucrados en la competencia, existen pocos resultados experimentales específicos, dado la dificultad metodológica de aislar la influencia de cada uno de los recursos. Gran parte del éxito competitivo de una especie depende de la proporción del total de recursos que ella pueda capturar en las primeras etapas de su crecimiento, antes incluso que se manifieste la competencia en la mezcla.
Cada tipo de cultivo lleva asociada una flora adventicia específica, debiéndose a que su período de establecimiento, sus ciclos biológicos y sus requisitos ecológicos son similares. En líneas generales éste es el factor más importante que condiciona la flora; dentro de una comunidad de plantas, no todas las especies tienen la misma importancia, normalmente existe 3 a 4 especies dominantes, que son las que originan la mayor parte de las reducciones de rendimiento del cultivo. También existen especies secundarias con una menor densidad y cobertura y por último algunas especies acompañantes cuya presencia es ocasional. La relación entre el número de malezas y el rendimiento del cultivo en la mayoría de los casos es de tipo sigmoidal, en donde unas pocas malezas no afectarán el rendimiento, mientras que la máxima reducción se consigue generalmente con una población de malezas (12 a 20 pl/m2), menor que la máxima posible de obtener.
Mientras el cultivo y las malezas son pequeñas no se producirá interferencia y el período comprendido entre emergencia hasta el momento en que no se producen efectos negativos en el rendimiento se denomina: Período Anterior a la Interferencia (PAI). Siendo el período máximo en el cual las malezas pueden ser toleradas por el cultivo sin afectar el rendimiento final. Desde emergencia hasta el punto después del cual las malezas no afectan mayormente el rendimiento de los cultivos se define como Período Total de Prevención de la Interferencia (PTPI). Este último concepto indica el período en que el cultivo debe estar libre de interferencia. El lapso de tiempo en que las malezas comienzan a interferir hasta aquel punto en que ya produjeron el máximo daño corresponde al Período Crítico de Interferencia (PCI), en la figura 1 se describen los diferentes períodos a nivel teórico de un cultivo anual tipo.
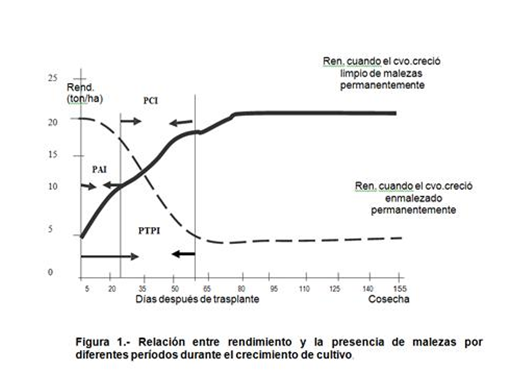
El PCI en los cultivos anuales se ha estimado que ocurre generalmente entre los 20 a 50 días después de emergencia. Este período coincide con una etapa de aumento en los requerimientos de los factores de producción. Las variaciones en la longitud o duración del PCI están normalmente relacionadas con el tipo de maleza, el cultivo, la región geográfica e incluso con la variedad. En el cuadro 4 se presenta el máximo período de interferencia tolerado sin afectar el rendimiento de los cultivos (PAI), recopilado por Zimdhal, 1980.
Cuadro 4.- Máximo período de interferencia tolerado sin afectar el rendimiento de diferentes cultivos.
|
Cultivo |
Duración del P.I. tolerado después de emergencia (Semanas) |
Malezas |
Localidad ylo país |
| Papa | 6 | Amaranthus retroflexus | Líbano |
| Papa | 4 | Eleusine indica, Panicum repens, Galinsoga parviflora, Polygonum aviculare |
Java, Indonesia |
| Zanahoria | 3 var. Kuroda | Cyperus rotundus | Brasil |
| Zanahoria | 5 a 7 var.Nantes | Mezcla de malezas anuales | Brasil |
| Repollo | 3 a 4 | Malezas anuales | Inglaterra |
| Repollo | 4 | Cyperus | Brasil |
| Cebolla | 4 | Mezcla de malezas anuales | Inglaterra |
| Cebolla | 5 | Amaranthus retroflexus, Polygonum persicaria | Wisconsin,EE.UU. |
| Cebolla | 12 | Amaranthus retroflexus, Gramíneas anuales | Nebraska, EE.UU. |
| Ajo | 3 | Cyperus rotundus | Brasil |
| Lechuga | 3 | Mezcla de malezas anuales | Inglaterra |
| Tomate trasplantado | 3 | Cyperus rotundus | Brasil |
Adaptado: Zimdhal, 1980.
- Solarización
Una de las alternativas no química, de manejo de malezas, es el uso de la radiación solar, en el suelo. El método consiste en cubrir el suelo húmedo con un plástico transparente que quede hermético, de modo que este se caliente por efecto de la energía solar. La humedad próxima a capacidad de campo en el suelo permite conducir el calor atrapado del sol hasta una profundidad de 20 cm. Casanova,S. et al., 2001 demostraron que en el sur de nuestro país, 30 días de solarización al suelo durante el mes de enero fueron suficientes para controlar semillas de malezas anuales, bajando la densidad de 450 pl/m2 a 3-4 pl/m2 ,en promedio, a la vez, el efecto se mantuvo persistente durante dos ciclos de cultivo de lechuga, indicando que llevando adelante este método una vez al año sería suficiente para controlar las malezas. Es un método inocuo para el medio ambiente, que debe tenerse en cuenta para todo lo referido a almácigos en tierra, preparación de suelo para almacigueras y manejo de las malezas bajo invernáculo; teniendo como desventajas el hecho que no controla propágulos vegetativos de malezas perennes y debe considerarse el alto costo del plástico así como la mano de obra para instalación.